Review of Related Literature of Diversity of Freshwater Fish
Abstract
Worldwide biodiversity assessments have mainly focused on species richness but little is known about the diverseness of species roles, i.eastward. functional diverseness, while this is a fundamental facet to understanding the consequences of global changes on the ecosystem services to human societies. Hither, nosotros report the world pattern of functional diversity of freshwater fish using a database encompassing morphological characteristics of more than ix,000 species. The Neotropical realm hosts more than than 75% of global functional diversity while other realms each host less than 25%. This discrepancy is mediated past loftier functional uniqueness in some diversified Neotropical fish orders. Surprisingly, functional diversity patterns were weakly related to functional vulnerability. In the Neotropics the loss of threatened species volition crusade a limited loss of functional diversity (<10%) whereas in the Nearctic and Palearctic realms, decline of the functional diversity will achieve 43% and 33%, respectively, conferring a high functional vulnerability to these realms. Conservation of the Neotropical fish diversity is a cardinal target to maintain globe fish functional variety, only this should not hide the pressing need to conserve the vulnerable fish faunas of the rest of the world, in which functional diversity is to a large extent supported by threatened species.
Introduction
Owing to the sixth mass extinction crisis1, jumbo efforts take been devoted to assessing biodiversity, merely to date, virtually worldwide biodiversity assessments have focused on the taxonomic component of biodiversity2,3,4,5. These worldwide assessments on taxonomic diversity take revealed that species richness varies strongly across the globe for most clades, e.g. birds5, plantshalf-dozen or fishesseven, with a decreasing richness gradient from the equator to the poles2. For the freshwater fish creature, the Neotropical realm (Due south and Central Americas) includes more 4,000 fish species, while the Palearctic realm (Eurasia, Middle-Eastward and North-Africa) contains fewer than 6008. As well this taxonomic facet, functional diversity (FD, i.e. the range of biological traitsnine) is a measure of the range of functions performed by organisms, and therefore a good mode to arroyo the role of biodiversity in sustaining ecosystem services9,10,11,12,13,xiv,15, likewise every bit the detrimental effects of homo disturbances on the erosion of biodiversity12. FD is expected to increment with taxonomic diversity (TD) because of the college probability of including a larger range of biological traits when more species are nowadaysxvi,17, but the influence of TD on FD is still unclear, as the lack of large-scale assessments of functional traits on a large range of species limits our capacity to run global scale studies on FDeighteen,19.
Hither, we appraise for the first time the FD of the native freshwater fish faunas over the half-dozen terrestrial biogeographic realms of the world. Functional diversity was measured as morphological diversitytwenty,21. Although these traits are not all direct linked to the actual roles played by fish in ecosystems (eastward.g. nutrient recycling22, trophic control of other taxa23), they remain informative to draw features related to nutrient acquisition and locomotion24,25,26, and remain the only traits measurable for near all fish species with a reasonable effort. We here measured x morphological traits on 9,170 fish species (see Supplementary Fig. S1 for details).
We quantified the extent to which FD, calculated as the book filled past the species in the multidimensional infinite defined by morphological traits, differs betwixt the biogeographic realms. And then, we tested the contribution of each fish order to each realm's functional diversity by measuring the functional uniqueness of each lodge within the realm (Funiq, i.east. proportion of the functional infinite of the realm filled only by this guild). Finally we assessed functional vulnerability to species loss equally the proportion of FD supported by species listed as 'threatened' past the IUCN27 (i.due east. condition is "critically endangered", "endangered" or "vulnerable"), and by species endemic to a single river drainage basin and not listed as "least concern" or "almost threatened" by the IUCN27,28.
Results and Discussion
Neotropics host more than three-quarters of the globe fish functional diversity
We report a threefold lower functional turnover (mean ± SD: 0.324 ± 0.206) than taxonomic turnover between realms (0.982 ± 0.022). Thus, despite strong species turnover between realms, the same core of biological attributes is shared between realms (Fig. 1B, Supplementary Fig. S2). Such a blueprint of functional nestedness betwixt realms hosting different suites of species has been reported for 2 other vertebrate taxa, passerine birds21 and coral reef fishnineteen.

(A) The six realms are displayed on the map. For each realm, the taxonomic (TD) and the functional diversity (FD) are represented equally a percentage of the world taxonomic or functional diversity. The number of species in each realm is given above the TD bar. (B) Functional infinite of the world freshwater fishes is the white area in the 2-dimensions space made by start and second principal component (PC) axes summarizing the five-dimensions functional space congenital based on 10 morphological traits (see other projections in Supplementary Fig. S2). The percentage of variance explained by each PC centrality is given in brackets (run into also Supplementary Figs S2 and S3). Fish functional diversity in each realm is illustrated past the polygon delimited by the black dashed line. The coloured polygons within it show the functional diversity of the about speciose fish orders. The map was generated using 'maps' and 'mapproj' libraries, available in R 3.0 (http://world wide web.r-project.org).
Besides this shared functional cadre, we show here a clear spatial discrepancy of fish FD among realms. The Neotropics host more than three-quarters of the earth FD (76.7%) whereas the two other tropical speciose realms, namely the Oriental and the Afrotropical realms, each hosts but one-fifth (17.8% and 20.eight%, respectively, Fig. 2). Hence, the twofold higher TD in Neotropics compared to the Afrotropics turns into a fourfold higher FD. A similar trend was observed between Neotropical and Oriental realms, confirming that the Neotropical fauna is functionally hyperdiverse (Fig. 2). Hence, the Neotropics, which stand for less than 15% of the world'southward continental surface, host more than 75% of the earth freshwater fish FD. In contrast, it is surprising to note that the FD of the Afrotropics is shut to that of the Palearctic realm, while the latter hosts less than one-half every bit many speciesviii. Similarly, the Australian realm hosts a FD shut to that of the Nearctic although information technology but hosts half as many species (Figs 1A and ii).

Functional diversity (FD) and taxonomic diverseness (TD) are expressed as a percentage of the world functional and taxonomic diversity of freshwater fish. The issue of species identity on functional diversity is represented by the mean (colored squares) and associated 95% confidence interval (black whiskers) computed on 999 values of a null model. This null model simulated realm assemblages using random sampling in the world pool of species, while conserving the taxonomic construction of the realm brute (i.eastward. number of species per order). Observed values of FD higher up the whiskers indicate a significant functional overdispersion, whereas values beneath the whiskers signal meaning functional clustering. The solid line represents the identity line FD = TD. Each color refers to ane realm.
The weak association we report between TD and FD suggests that the multifariousness of species traits present in a realm is non a random subset of the pool of traits values at the world scale. Indeed, we institute that the Neotropical fish animate being has a significantly higher FD than expected given its TD (Standardized Effect Size (SES) = 2.21, P > 0.99) whereas the FD of Afrotropical (SES = −2.46, P < 0.01) and Oriental realms (SES = −2.l, P < 0.01) were significantly lower than expected (Fig. 2). FD did not differ from random expectation for Australian brute (SES = −0.79, P = 0.22) whereas a significantly lower FD was reported for Nearctic (SES = −1.32, P < 0.05) and Palearctic realms (SES = −ane.54, P < 0.05). The functional overdispersion of the Neotropical fish animate being could be due to (i) either a few functionally diversified orders that make full most of the functional space (the remaining functionally poor orders being nested in the functionally rich orders), or (ii) a high functional dissimilarity between orders, i.e. each gild filled a unique part of the realm FD. On the other paw, the functional clustering of the Afrotropical and Oriental realms could be driven past (i) a low functional diversity of all orders and/or (2) a high functional similarity (i.eastward. redundancy) between orders.
Differences in functional diversity are driven past the functional uniqueness of a few fish taxa
Although the FD of orders at the global scale was correlated to their TD (Spearman'due south rank correlation rho = 0.31, n = 46, P < 0.05), the functional uniqueness (Funiq) of orders was independent of order TD (Spearman's rank correlation rho = −0.04, due north = 46, P = 0.76). Indeed, the realm FD results non merely from the global FD of each order (Fig. one), merely also from the degree of functional uniqueness between orders (Fig. 3). The functional clustering for all realms except the Neotropics is thus due to the depression functional uniqueness of their near speciose orders (Fig. iii). This is particularly striking for the Afrotropics, where the two most speciose orders (Perciforms and Cypriniforms) that together account for more than 60% of the Afrotropical species richness, contribute marginally to the realm FD (five.2 and ii.vi%, respectively) because of their low Funiq (26.2 and 0.4%, respectively, Fig. 3). This was verified for the Perciforms, despite the recent and intense radiation of cichlids (the nigh species rich family of Perciforms in the Afrotropics) promoted past the contempo opening of the rift lakes (e.g. Malawi, Tanganyika, Victoria) that produced hundreds of phylogenetically closely related species29. Although the cracking lakes cichlids radiations have generated various morphological adaptations29, the resulting morphological features did not strongly differ from those already experienced within the African fish fauna, hence explaining the low functional uniqueness of the Afrotropical fish orders.

The functional diversity (FD) of each fish order in each realm is given every bit the percentage of the world FD (bars). The part of FD of each guild non-filled by any other order in the realm (Functional uniqueness, Funiq) is the ratio between the grey part and the FD of each society. Annotation that the FD scale differs between realms. The orders are sorted by decreasing TD in each realm and the black dots indicate the percentage of the realm TD represented by each society. Only the 13 nearly speciose orders in the earth are represented (Acipens: Acipenseriforms; Atheri: Atheriforms; Belonif: Beloniforms; Characi: Characiforms; Clupei: Clupeiforms; Cyprini: Cypriniforms; Cyprino: Cyprinodontiforms; Gymnoti: Gymnotiforms; Osteogl: Osteoglossiforms; Osmerif: Osmeriforms; Percifo: Perciforms; Salmoni: Salmoniforms; Silurif: Siluriforms).
In contrast, the Neotropical fish animate being hosts species with unique functional attributes such as i) extremely elongated fish with a large last mouth and a loftier caudal peduncle throttling, corresponding to mobile surface predators such as some of the Beloniforms, or 2) dorso-ventraly flattened fishes, with a ventral mouth located below the caput and a small caudal peduncle throttling, mainly corresponding to benthic algae browsers with express pond efficiency such every bit some Loricariid species (run across Supplementary Fig. S3). The Loricariids belong to the Siluriforms that is the most speciose order in the Neotropics and they strongly contribute to the high FD of the realm due to their high Funiq (62.5%, Fig. 3). Indeed Neotropical Siluriforms have an impressive trunk size range (from less than v cm to more than 200 cm), a broad span of diet (from algae browsing in the Loricaridae, to ichthyophagy in the Pimelodidae, and even parasitism in the Trichomycteridae, see Supplementary Fig. S3), and of habitat associated with diverse body shapes from flattened to extremely elongated30. Such a high FD of Siluriforms is not observed in the other realms, explaining their depression functional uniqueness (6.6% in Oriental, 7.iv% in Nearctic and 7.1% in Palearctic realms, Fig. 3) despite their contribution to realm TD (e.g. 373 Siluriforms species in the Afrotropical realm).
Functional vulnerability peaks in the Nearctic and Palearctic realms, despite their low functional multifariousness
The loss of FD through species extinction is expected to be marked when threatened species support unique functions, every bit shown in alpine plants, tropical copse, coral reef fishes and bird regional assemblages19,31,32. For freshwater fish, the vulnerability of FD to the extinction of threatened species differed amongst realms by a sixfold gene (Fig. 4), and was neither correlated to the realm FD (Spearman'due south rank correlation rho = −0.43, P = 0.42) nor to the number of threatened species (Spearman'southward rank correlation rho = −0.08, P = 0.92). The loss of FD due to the simulated extinction of threatened species was marked in the Nearctic and Palearctic realms (43.6 and 33.5%, respectively, Fig. 4), while it was low (from seven.47 to fourteen.1%) in the Afrotropical, Australian, Neotropical and Oriental realms. Inside this last group, the Neotropics are characterized by a vulnerability of FD to species loss lower than expected given the number of threatened species (SES = −2.32, P < 0.001, Table 1). In contrast, in the Nearctic realm the vulnerability of FD was significantly higher than expected given the number of threatened species (SES = iii.47, P > 0.999, Table ane). A similar trend although not significant was found in the Palearctic realm (SES = 0.84, P = 0.66, Table 1). While being the least functionally diverse, the Nearctic and Palearctic realms are the near vulnerable to species loss (Table ane). Although there is a large number of threatened species (743 species accounting for 31% of the Afrotropical fish beast), the Afrotropical vulnerability is low whereas in the Nearctic and Palearctic realms, fewer species are threatened (147 and 317 species, respectively), but they differ functionally from the rest of the fauna (east.g. sturgeons and eels) and thus pb to a high vulnerability of the functional multifariousness of this realm. As for alpine plants, tropical copse and coral reef fishes32, the threatened species back up vulnerable functions, testifying that the imminent extinction of a few endangered species will cause a marked turn down of functional diversity across the world faunas.

(A) Taxonomic vulnerability. Bars represent the proportion of threatened species (i.east. species with an IUCN status "CR", "EN" or "VU" or species occurring in only one drainage basin beyond the earth which IUCN status was non "Well-nigh threatened" nor "Least concern"). The hatched part of each bar represents the taxonomic vulnerability due merely to the virtually threatened species (i.e. the species listed as "CR", "EN" or "VU" by the IUCN). (B) Functional vulnerability. Bars represent the loss of functional diverseness (FD) if threatened species become extinct. The hatched part of each bar represents the functional vulnerability due simply to the almost threatened species (i.e. the species listed as "CR", "EN" or "VU" by the IUCN).
In addition, when considering but the well-nigh threatened species, i.eastward. those listed as critically endangered (CR), endangered (EN) or vulnerable (VU) by the IUCN, we institute that these species account for virtually of the functional vulnerability in the Nearctic (94.4%) and Oriental (70.5%) realms and to a lower proportion in the Palearctic (51.four%) and fifty-fifty less than 25% in the 3 others realms. In those concluding realms, nigh of the vulnerability is supported by endemic species that are potentially threatened although being not evaluated yet (Fig. 4, Table 1). Extending the IUCN evaluation of threats to the world fish brute is therefore a prerequisite to determine the role played by the most threatened species in the maintenance of the functional diversity over more than half of the world continental areas.
Conclusion
Assessing functional diversity and its determinants on large scale is pivotal in biodiversity mapping11. Indeed, the species number is not a good surrogate for functional diversity as illustrated by the contrasting FD observed hither betwixt the two most speciose realms, namely the Neotropics and the Afrotropics. Moreover, FD does not match with functional vulnerability because of the particular functional attributes of threatened species in the Palearctic and Nearctic realms. These mismatches between TD and FD and between FD and functional vulnerability stress the need to consider multiple biodiversity facets in efficient conservation planning. More than precisely, time to come conservation efforts should focus on the vulnerable realms, namely the Nearctic and Palearctic, just also in the Neotropics to protect the habitats hosting the functionally about original species across the world. Such conservation actions should exist taken without delay, as the vulnerable functions are overrepresented among the already-listed threatened species. In improver, as the endemic species which status are still unknown also contribute to a high proportion of functional diversity over more than one-half of the world continental surface, studies at the species level are urgently needed to evaluate the threats they are facing. Because more than specifically freshwater fish, that constitute an emblematic case of massive introductions of non-native species3, information technology is urgent to assess whether the patterns reported here for historical native fish faunas have been blurred by the biotic exchanges betwixt continents for the last centuries. Towards constructive assessments of biodiversity-ecosystem services relationships, the big-scale patterns reported here should exist completed by local assessments of the key ecological roles played by fish in aquatic ecosystems.
Methods
Fish occurrence database
We considered the half dozen terrestrial biogeographic realms (Afrotropical, Australian (including Oceania), Nearctic, Neotropical, Oriental and Palearctic) commonly used for freshwater fish8,33,34,35. The number and identity of the species occurring in each realm was obtained using the database on freshwater fish occurrences34 that contains ix,170 freshwater fish species out of the ca. xiii,000 strictly freshwater fish species describedxxx, and hence covers 77% of the documented earth freshwater fish fauna.
Fish pictures collection
Nosotros hither developed the most comprehensive fish functional database existing to engagement. It encompasses ix,170 freshwater fish species out of the ca. xiii,000 described strictly freshwater fish species (i.due east. 77% of the globe freshwater fish fauna). We ran an extensive literature review to collect at least one lateral view moving picture of each species, which were used to measure morphological characteristics (Supplementary Fig. S1a). Some of the pictures were taken on the field by the authors. Fish captures were led in accordance with laws and guidelines apropos live animals, and all the experiments were canonical past the Direction of Environment of the French ministry of environment (DEAL), the French Guiana National Park (Parc Amazonien de Guyane), and the research program LABEX CEBA (ANR-10-LABX-25-01).
For all the species nowadays in the taxonomic database nosotros aimed at measuring 10 functional traits describing species strategies for food conquering and locomotion (Supplementary Fig. S1 and24). Fish size was described using the log-transformed maximum body length (values taken from FishBase36). In improver to size, nine morphological traits were measured on side view pictures (Supplementary Fig. S1a) collected during an extensive literature review from more than 200 scientific literature sources including peer-reviewed articles, books and scientific websites. Nosotros collected at least one picture (validated photograph or scientific cartoon) per species. Merely adept quality pictures and scientific side view drawings of entire developed animals were kept. Juveniles were not considered as morphological changes can occur during ontogeny. In the event of sexual dimorphism, we only considered male morphology, as female pictures are scarce for almost species (peculiarly for Perciforms and Cyprinodontiforms). Using lateral view pictures did non let to collect every bit much external morphological data as fresh animals (due east.k. oral gape surface, body transversal shape). It would nevertheless exist very demanding to collect fresh or museum specimens for more than 9,000 species, and using lateral view pictures was the most efficient way to collect morphological measures for more than 70% of the globe freshwater fish animal.
Functional traits
For each specimen, 11 morphological measurements were recorded (come across Supplementary Fig. S1a) using ImageJ software (http://rsb.info.nih.gov/ij/index.html) and were then used to compute nine unitless ratios describing the morphology of the fish caput (including mouth and eye), body, pectoral and caudal fins (Supplementary Fig. S1b). The 10 functional traits (9 unitless ratios and size) selected are commonly used in cess of fish functional multifariousness (e.yard.25,26,37). Complementary functional traits (e.g. gut length, oral gape surface area and shape, fecundity) were not included because they are only available for a few species. The quality of the pictures did not allow measurement of all 11 morphological traits in all species, but more than 80% of the species were functionally characterized within each taxonomic order. All nine morphological traits were measured for almost two-thirds of the resulting morphological database (6,030 species) and overall xviii% of the morphological measurements were not obtained.
Some species have unusual morphologies (species without tail, flatfishes) that forbid from measuring some morphological traits. We thus defined rules for these few exceptions as in26: (i) for species with no visible caudal fin (east.g. Sternopygidae, Anguilidae, Plotosidae), Caudal Peduncle Throttling was set up to 1 (assuming CFd = CPd, meet Supplementary Fig. S1b), (two) for the species with the mouth positioned under the body (e.g. Loricaridae, some Balitoridae such every bit Gastromyzon) mouth vertical position (Mo) was set to 0, (iii) for the species without pectoral fins (e.g. Synbranchiforms and some Anguiliforms) Pectoral fin vertical position (PFl/Bd) was set up to 0.; iv) for flatfishes Bd was the trunk width as the fish lies on 1 side of its body. We hence assumed that Pleuronectiforms are functionally closer to dorso-ventrally flattened fishes (e.thousand. Gastromyzon) than to laterally compressed fishes (eastward.g. Symphysodon).
Other traits, such as ecological, behavioural or physiological traits were not considered in this study, because there is currently no database available at the world scale.
Functional space
The 10 functional traits were ordered in a multidimensional functional space by means of Chief Components Analysis using a regularized algorithm designed for ordination analysis to handle the missing values38. The start v axes account for eighty.5% of the full variance (each selected centrality had an eigenvalue >1) (Supplementary Fig. S4) and were retained to build a 5-dimensional functional infinite. We assessed the robustness of our findings using sensitivity procedure. Nosotros tested the outcome of trait identity on the functional distance between species past rerunning all analyses using all combinations of nine functional traits out of x. The results were inappreciably affected by this procedure (Mantel tests r > 0.900, P < 0.001). Thus, our findings are non affected by functional trait selection.
Threatened species
Two types of threatened species were considered, the "most threatened" and the "potentially threatened". The "most threatened" species were identified as the species listed as critically endangered ('CR'), endangered ('EN') and vulnerable ('VU') in the well-nigh recent IUCN Crimson Listing assessment27. The IUCN Cherry-red List of threatened species although existence the nigh objective and authoritative organisation for classifying species in terms of the risk of extinction at the global scale, remains affected by spatial disparities in species evaluation. To deal with incompleteness of the IUCN assessment (52.v% of the species occurring in our database have non been IUCN evaluated, see Supplementary Table S1), strictly endemic species (species occurring in only i drainage basin across the globe28,30,34) which IUCN status was not "well-nigh threatened (NT)" nor "least concern (LC)" were considered equally "potentially threatened". Those species with limited spatial distribution take been recognized as prone to extinction following human disturbance4,28,39. Thus, a total of iii,443 (37.5%) species out of the ix,170 species were considered equally threatened.
Biodiversity indices
Taxonomic diversity (TD) was calculated as the number of species in each realm (or in each order). Functional diversity (FD) was assessed with the functional richness alphabetize (FRicforty), computed as the volume of the minimum convex hull that includes all the species in the 5-dimensional functional space. The higher the FD, the higher the richness of combinations of functional trait values in the species pool considered. FD and TD indices were computed for all species in each realm, and for all orders within each realm.
Statistical analyses
To exam whether observed functional multifariousness of a realm was significantly different from functional multifariousness of a random subset of species, we used null models based on randomization of species pools. To simulate a realistic puddle of species in each realm, the number of species per taxonomic gild was kept constant in the random selection procedure (999 iterations). The standardized effect size (SES) was used to measure the deviation between observed values (obs) and null expectation (rand): SES = (FDobs − mean (FDrand))/sd (FDrand). The significance of the departure from nil expectations was tested using a 2-tailed test (∝ < 0.05). When FDobs is lower than 97.5% of FDrand values, the assemblage is considered as functionally amassed. When FDobs is greater than 97.5% of FDrand values, the assemblage is considered as functionally overdispersed.
The dissimilarity in FD and TD between realms was measured as taxonomic and functional turnover41. Taxonomic turnover measures the replacement of species between two assemblages independently of the departure in species richness41. Similarly, functional turnover measures the replacement of functional book independently of the difference in functional richness. Taxonomic and functional turnover were calculated using the same formula: 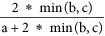 , where a is the number of species (or the volume) shared past the two assemblages and b and c the number of unique species (or the unique volume) hosted by the ii assemblages41,42. Both taxonomic and functional turnover varies from 0, when the two assemblages are completely similar, to 1 when the 2 assemblages shared no species (taxonomic) or no portion of the functional infinite.
, where a is the number of species (or the volume) shared past the two assemblages and b and c the number of unique species (or the unique volume) hosted by the ii assemblages41,42. Both taxonomic and functional turnover varies from 0, when the two assemblages are completely similar, to 1 when the 2 assemblages shared no species (taxonomic) or no portion of the functional infinite.
The functional uniqueness of each fish society o in each realm r was computed as the proportion of the functional space filled merely by the order considered: 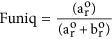 , where a r o is the volume filled only by focal gild o and b r is the book shared with all other orders present in realm r. Taxonomic and functional turnover, and Funiq were computed using the 'functional.beta.core' function from the 'betapart' R package43.
, where a r o is the volume filled only by focal gild o and b r is the book shared with all other orders present in realm r. Taxonomic and functional turnover, and Funiq were computed using the 'functional.beta.core' function from the 'betapart' R package43.
The functional vulnerability (FV) to threatened species loss in each realm was computed as the proportion of the FD of the realm that will remain if all threatened species got extinct: FVobs = (FDobs − FDnon threatened)/FDobs. Functional vulnerability is null when species not threatened contribute fully to the FD of the realm and tends to the 100% if all the most extreme combinations of traits are supported merely past threatened species.
To test the significance of the observed functional vulnerability given the number of threatened species we designed a null model based on a random choice of the species expected to exist lost (999 iterations). The standardized effect size (SES) was used to measure the difference betwixt predicted FD loss values due to extinction of threatened species (FVobs) and nothing expectation of FD loss (rand): SES = (FVobs − mean(FVrand))/sd (FVrand). The significance of the divergence from goose egg expectations was tested using a 2-tailed test (∝ < 0.05). A p-value lower than 0.025 indicates that threatened species are functionally distinct and over-contribute to FD whereas a p-value higher than 0.975 indicates that threatened species are redundant with other species of the realm FD.
We then ran the aforementioned analyses considering only the most threatened species, i.e. those listed as "critically endangered", "endangered" or "vulnerable" by the IUCN27.
All statistical analyses were performed with R software version three.044.
Boosted Information
How to cite this article: Toussaint, A. et al. Global functional diversity of freshwater fish is full-bodied in the Neotropics while functional vulnerability is widespread. Sci. Rep. 6, 22125; doi: 10.1038/srep22125 (2016).
References
-
Barnosky, A. D. et al. Has the Earth'due south 6th mass extinction already arrived? Nature 471, 51–57 (2011).
-
Gaston, K. J. Global patterns in biodiversity. Nature 405, 220–227 (2000).
-
Leprieur, F., Beauchard, O., Blanchet, South., Oberdorff, T. & Brosse, South. Fish invasions in the earth's river systems: when natural processes are blurred by homo activities. PLoS Biol. half dozen, e28 (2008).
-
Pimm, Southward. L. et al. The biodiversity of species and their rates of extinction, distribution, and protection. Science 344, 1246752 (2014).
-
Jetz, W., Sekercioglu, C. H. & Böhning-Gaese, K. The Worldwide Variation in Avian Clutch Size beyond Species and Space. PLoS Biol. 6, e303 (2008).
-
Devictor, V. et al. Spatial mismatch and congruence betwixt taxonomic, phylogenetic and functional diverseness: the demand for integrative conservation strategies in a irresolute world. Ecol. Lett. xiii, 1030–1040 (2010).
-
Oberdorff, T. et al. Global and Regional Patterns in Riverine Fish Species Richness: A Review. Int. J. Ecol. 2011, ane–12 (2011).
-
Lévêque, C., Oberdorff, T., Paugy, D., Stiassny, 1000. L. J. & Tedesco, P. A. Global diversity of fish (Pisces) in freshwater. Hydrobiologia 595, 545–567 (2007).
-
Violle, C. et al. Let the concept of trait exist functional! Oikos 116, 882–892 (2007).
-
Cadotte, M. W., Carscadden, K. & Mirotchnick, North. Beyond species: functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 48, 1079–1087 (2011).
-
Mouillot, D., Villéger, S., Scherer-Lorenzen, M. & Mason, N. W. H. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS One 6, e17476 (2011).
-
Cardinale, B. J. et al. Biodiversity loss and its touch on humanity. Nature 486, 59–67 (2012).
-
Srivastava, D. Due south., Cadotte, K. Due west., MacDonald, A. A. M., Marushia, R. K. & Mirotchnick, N. Phylogenetic diverseness and the operation of ecosystems. Ecol. Lett. 15, 637–648 (2012).
-
Díaz, S. et al. Incorporating institute functional diversity effects in ecosystem service assessments. Proc. Natl. Acad. Sci. U.s.a. 104, 20684–20689 (2007).
-
D'agata, S. et al. Homo-mediated loss of phylogenetic and functional diverseness in coral reef fishes. Curr. Biol. 24, 555–560 (2014).
-
Tilman, D., Knops, J., Wedin, D. & Reich, P. The Influence of Functional Diversity and Composition on Ecosystem Processes. Science. 277, 1300–1302 (1997).
-
Cornwell, West. Thousand., Schwilk, L. D. W. & Ackerly, D. D. A trait-based test for habitat filtering: convex hull volume. Ecology 87, 1465–1471 (2006).
-
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J. & Kattge, J. The emergence and promise of functional biogeography. Proc. Natl. Acad. Sci. USA 111, 13690–13696 (2014).
-
Mouillot, D. et al. Functional over-redundancy and high functional vulnerability in global fish faunas on tropical reefs. Proc. Natl. Acad. Sci. USA 111, 13757–13762 (2014).
-
Azzurro, East. et al. External morphology explains the success of biological invasions. Ecol. Lett. 17, 1455–1463 (2014).
-
Ricklefs, R. E. Species richness and morphological diversity of passerine birds. Proc. Natl. Acad. Sci. United states 109, 14482–14487 (2012).
-
McIntyre, P. B. et al. Fish distributions and nutrient cycling in streams: Tin fish create biogeochemical hotspots? Ecology 89, 2335–2346 (2008).
-
Brandl, South. J. & Bellwood, D. R. Individual-based analyses reveal limited functional overlap in a coral reef fish community. J. Anim. Ecol. 83, 661–670 (2014).
-
Winemiller, Thousand. Ecomorphological diversification in lowland freshwater fish assemblages from five biotic regions. Ecol. Monogr. 61, 343–365 (1991).
-
Bellwood, D. R., Goatley, C. H. R., Brandl, S. J. & Bellwood, O. Fifty million years of herbivory on coral reefs: fossils, fish and functional innovations. Proc. Biol. Sci. 281, 20133046 (2014).
-
Villéger, South., Ramos Miranda, J., Flores Hernández, D. & Mouillot, D. Contrasting changes in taxonomic vs. functional multifariousness of tropical fish communities afterwards habitat degradation. Ecol. Appl. xx, 1512–1522 (2010).
-
IUCN. The IUCN Carmine Listing of Threatened Species. Version 2015-iv. http://www.iucnredlist.org. Downloaded on 19 November 2015 (2015).
-
Tedesco, P. A. et al. Patterns and processes of global riverine fish endemism. Glob. Ecol. Biogeogr. 21, 977–987 (2012).
-
Albertson, R. C., Markert, J. A., Danley, P. D. & Kocher, T. D. Phylogeny of a rapidly evolving clade: the cichlid fishes of Lake Republic of malaŵi, Due east Africa. Proc. Natl. Acad. Sci. USA 96, 5107–5110 (1999).
-
Nelson, J. S. Fishes of the Earth. (Wiley, 2006).
-
Thuiller, W. et al. The European functional tree of bird life in the confront of global change. Nat. Commun. 5, 3138 (2014).
-
Mouillot, D. et al. Rare species support vulnerable functions in high-diversity ecosystems. PLoS biol. xi, e1001569 (2013)
-
Villéger, South., Blanchet, S., Beauchard, O., Oberdorff, T. & Brosse, S. Homogenization patterns of the world's freshwater fish faunas. Proc. Natl. Acad. Sci. USA 108, 18003–18008 (2011).
-
Brosse, S. et al. Fish-SPRICH: a database of freshwater fish species richness throughout the World. Hydrobiologia 700, 343–349 (2013).
-
Leprieur, F. et al. Division global patterns of freshwater fish beta diversity reveals contrasting signatures of by climate changes. Ecol. Lett. 14, 325–334 (2011).
-
Froese, R. & Pauly, D. Fishbase. World Wilde Web electronic publication. Available at: http://www.fishbase.org (accessed on 8 May 2013) (2013).
-
Pease, A. A., Taylor, J. M., Winemiller, Thousand. O. & King, R. South. Ecoregional, catchment, and reach-scale environmental factors shape functional-trait structure of stream fish assemblages. Hydrobiologia 753, 265–283 (2015).
-
Josse, J. & Husson, F. Handling missing values in exploratory multivariate information assay methods. J. Soc. Fr. Statistique. 153, 79–99 (2012).
-
Vitousek, P. M. Human being Domination of Globe's Ecosystems. Science 277, 494–499 (1997).
-
Villéger, South., Mason, North. Due west. H. & Mouillot, D. New multidimensional functional variety indices for a multifaceted framework in functional ecology. Environmental 89, 2290–2301 (2008).
-
Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 21, 1223–1232 (2012).
-
Villéger, S. & Brosse, South. Measuring changes in taxonomic dissimilarity following species introductions and extirpations. Ecol. Indic. eighteen, 552–558 (2012).
-
Baselga, A. & Orme, C. D. L. Betapart: an R parcel for the report of beta diversity. Methods Ecol. Evol. three, 808–812 (2012).
-
R Development Core Team. R: A Language and Environment for Statistical Calculating. (2014). at http://www.r-project.org/.
Acknowledgements
We thank F. Leprieur and C. Violle for their comments that helped us to improve a kickoff version of this manuscript and Peter Winterton for correcting the English. This work was supported by the European union BioFresh Project (7th Framework European Program, Contract No. 226874) and by the 'Laboratoires d'Excellence' (LABEX) entitled TULIP (ANR-ten-LABX-41) and CEBA (ANR-10-LABX-25-01). A.T. was supported by the French National Middle of Scientific Research (CNRS, Contract No. 360776).
Writer data
Affiliations
Contributions
A.T., Due south.B. and Southward.V. designed the study, discussed the results and wrote the manuscript. N.C. and A.T. built the database and A.T. performed the analyses.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary data
Rights and permissions
This work is licensed nether a Creative Commons Attribution 4.0 International License. The images or other third party material in this commodity are included in the article's Artistic Commons license, unless indicated otherwise in the credit line; if the fabric is not included under the Artistic Commons license, users will need to obtain permission from the license holder to reproduce the cloth. To view a copy of this license, visit http://creativecommons.org/licenses/by/four.0/
Reprints and Permissions
About this commodity

Cite this commodity
Toussaint, A., Charpin, North., Brosse, S. et al. Global functional diversity of freshwater fish is concentrated in the Neotropics while functional vulnerability is widespread. Sci Rep 6, 22125 (2016). https://doi.org/10.1038/srep22125
-
Received:
-
Accepted:
-
Published:
-
DOI : https://doi.org/x.1038/srep22125
Further reading
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag information technology as inappropriate.
Source: https://www.nature.com/articles/srep22125
0 Response to "Review of Related Literature of Diversity of Freshwater Fish"
Post a Comment